科学传播
大冰川中的小精灵—青藏高原冰川微生物
地球母亲不仅孕育了丰富多样的宏观生命,还滋养了几乎无处不在的微观生命,即微生物。微生物是地球上最古老的生物类群,在生物界占据举足轻重的地位。主要的微生物类群包括:原核生物细菌和古菌,真核生物真菌和藻类,以及无细胞结构不能独立生活的病毒等。微生物具有形态微小、代谢高效、分布广泛和适应性强等特点,通过主导物质的分解、转化过程成为生物地球化学循环中不可缺少的成员,也通过在冰川、冻土、热泉、盐湖等极端环境中的适应性生存定义了生命的边界。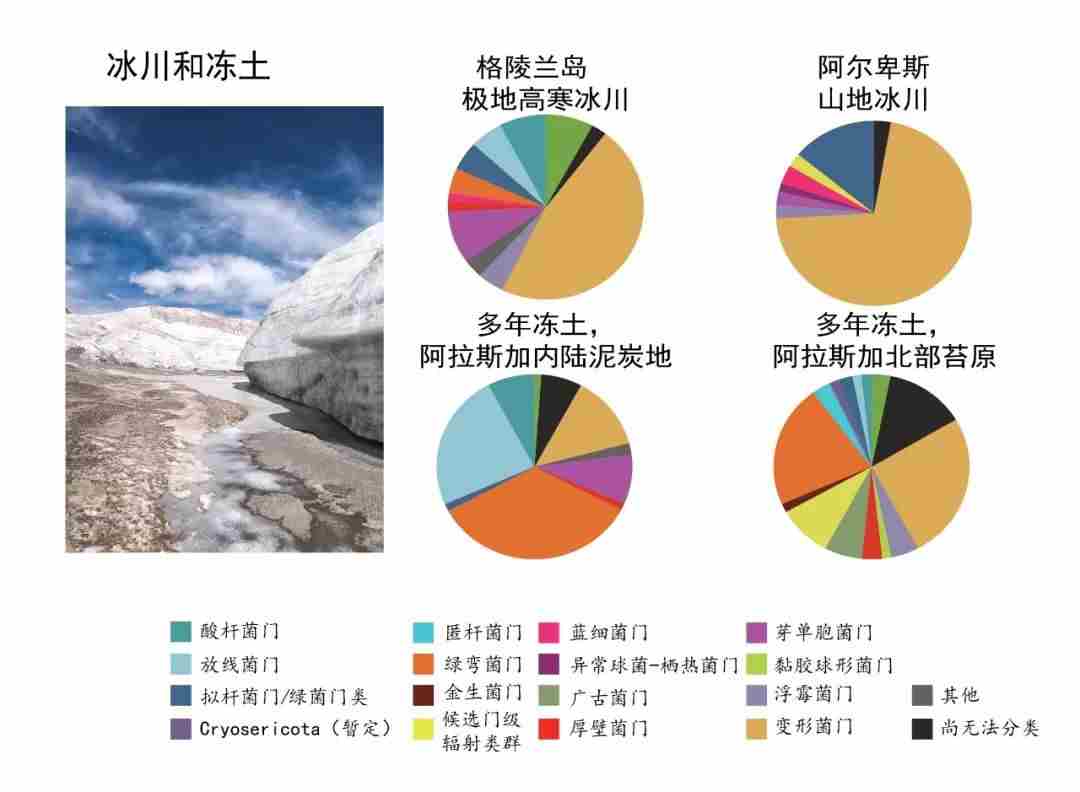
图1. 冰川冻土极端环境中不同的微生物定义了生命的边界,图源:改自文献[3]。
冰川覆盖约10%地球陆地面积,贮存了冰川数百年数万年甚至数百万年以来的各种微生物,代表了地球未知基因库的很大一部分。在冰川形成时,来自周边生态系统沉降至冰川表面的微生物被一起封存;当冰川随气候变暖消融时,这些微生物也会随冰川融水再次进入下游生态系统。根据每升冰川融水中的微生物数量及冰川消融量,可估算每年有1017-1021个微生物随着冰川融化被释放,进而改变下游生态系统的微生物群落和生物地球化学循环过程。

图2. 冰川微生物向下游径流和湖泊释放示意图(左)及青藏高原枪勇冰川及冰前湖(右)。冰川是下游水生生态系统重要的微生物物种库,向下游径流和湖泊贡献了高达73%-82%的物种;冰川径流是冰川来源微生物的接受体和载体,径流越短,冰川来源的物种对湖泊微生物的影响越大;微生物在冰川补给水文连续体中的迁移模式受质量效应的影响,并受到各生境局部环境的选择作用,图源:第二次科考队。
巍峨雄伟的青藏高原上分布着南北极以外地球上面积最大的冰川,蜿蜒连绵,构成雪白纯洁晶莹透剔的冰雪世界景观。青藏高原冰川是亚洲水塔的固体水库,储存着南北极外最大量的固态淡水;青藏高原冰川也是过去地质和历史时期微生物的天然存储库,封存了季风环流搬运、西风环流搬运和局地生长的不同种类微生物。青藏高原是西风带与亚洲季风环流系统交汇区,所以季风环流搬运的微生物来自海洋及沿途不同生态系统的微生物,西风环流搬运的微生物则源于欧洲及中亚等地区。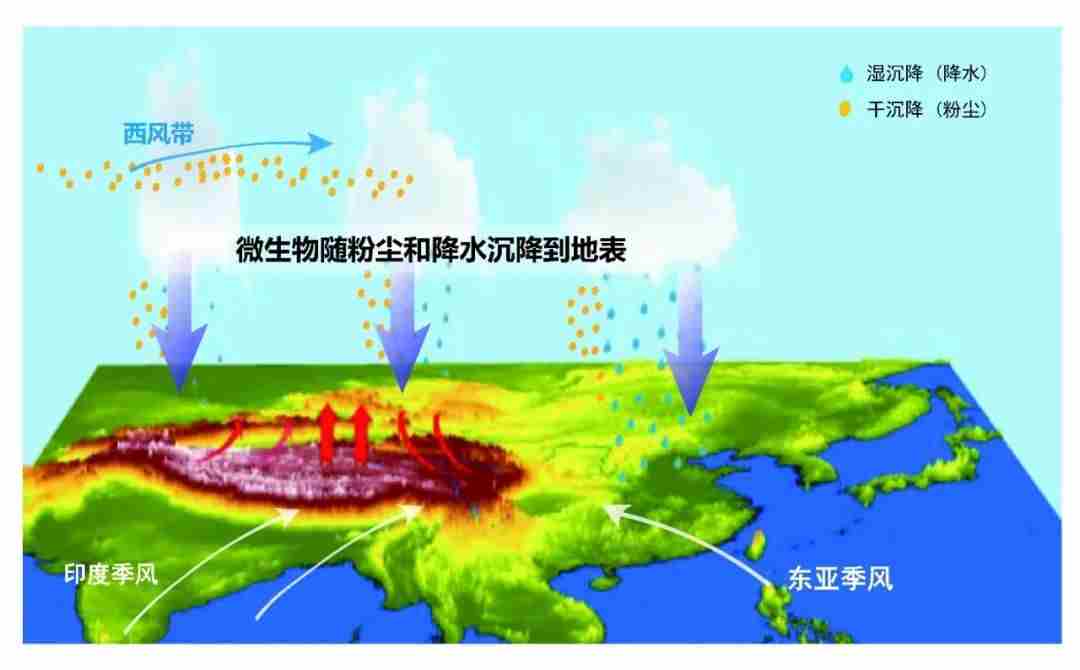
图3. 西风季风带来不同生态系统的微生物沉降到青藏高原,图源:第二次科考队。
微生物沉降到冰川,是冰川极端环境中主要生命形态,驱动着冰川生态系统的碳氮硫元素循环。冰川景观看似单一,其实有不同的小生境,生存着不同种类的微生物。低温、强紫外辐射等极端环境条件塑造了独特的冰川微生物类群。
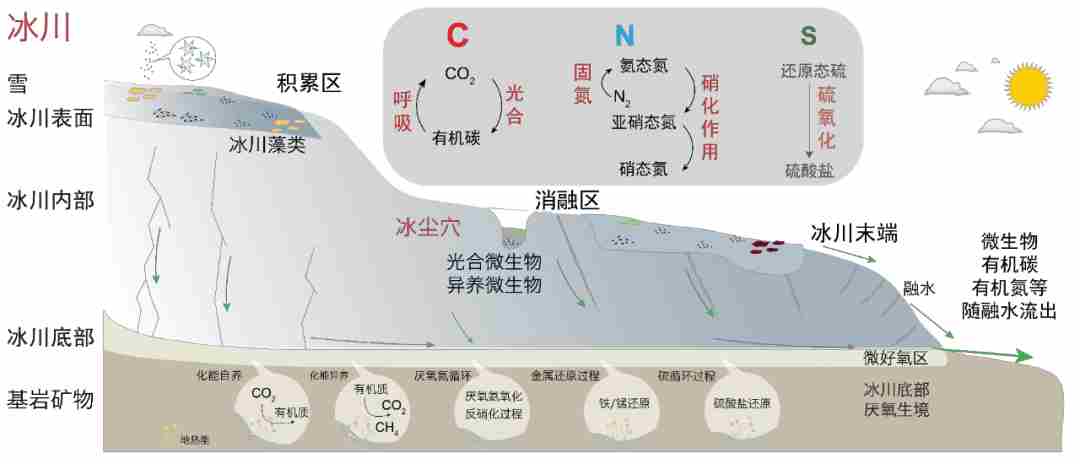
图4. 冰川生态系统示意图,图源:第二次科考队。

图5.珠穆朗玛峰东绒布冰川:是建立青藏高原冰川微生物基因组和基因数据集的典型冰川,图源:第二次科考队。
全球变暖导致中低纬度冰川快速消融,适应冰川环境的特有微生物不断消失,大量微生物在冰川消融时随冰川融水向下游释放。青藏高原正在遭遇高于全球两倍的增温,冰川快速消融,我们亟待了解青藏高原冰川中特有微生物如何向下游释放。第二次青藏高原综合科学考察研究的队员们针对气候变化下冰川微生物多样性快速损失的现状,系统开展青藏高原冰川微生物基因组及基因数据系统收集与研究工作,以摸清我国青藏高原冰川的生物多样性家底,明确生物多样性热点区域,揭示冰川微生物多样性与气候环境的关系,从而实现有有针对性的生物多样性保护,更好地发挥青藏高原的生态屏障功能。

图6.青藏高原唐古拉龙匣宰陇巴冰川表面的冰尘穴是冰川表面微生物数量和活性很高的生境,图源:第二次科考队。
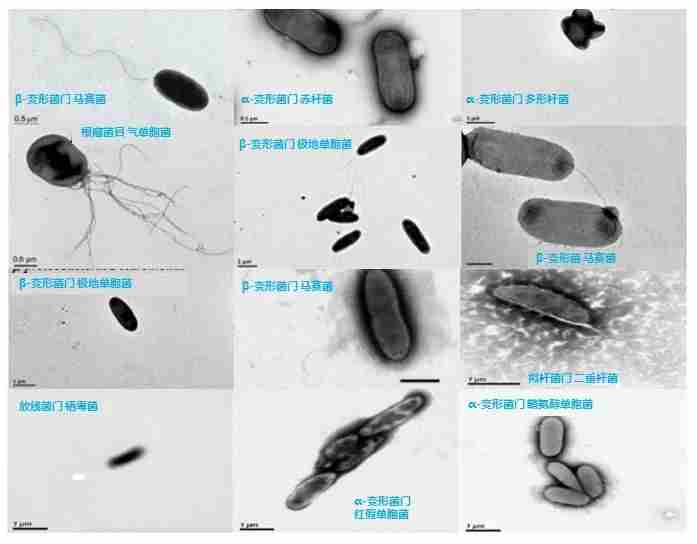
图7. 从青藏高原冰川中分离培养的细菌新种,图源:第二次科考队。
冰川—延展微生物物种认知边界的魔法世界
科考队员从青藏高原21条冰川采集了85个雪、冰和冰尘样品进行宏基因组测序,加上分离自青藏高原冰川的883个细菌的基因组序列,构建了青藏高原冰川微生物基因组与基因数据集,这也是全球首个冰川微生物数据集。获得了3241个冰川微生物(细菌和古菌)基因组,分属30个门,69个纲,12个目,22个科,475个属和968个种。
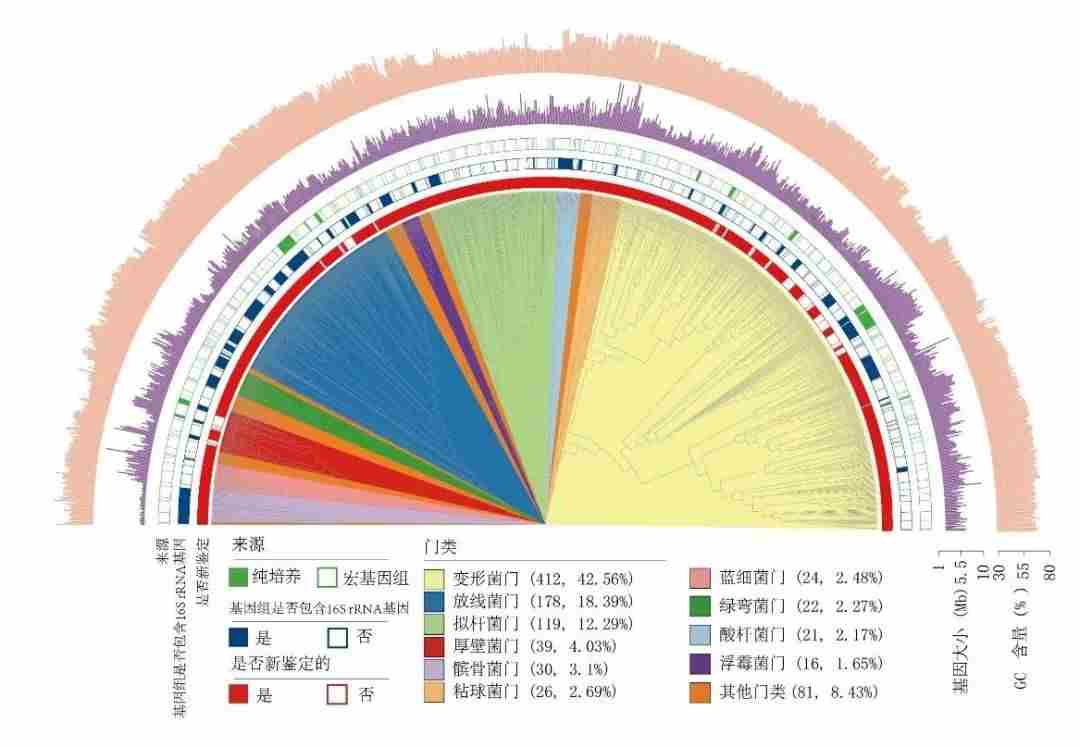
图8.获取的青藏高原冰川微生物968种细菌代表基因组系统发育树,图源:第二次科考队。
科考发现,青藏高原冰川蕴藏大量从未被发现过的微生物类群:与极地海洋和地球微生物数据库中的基因组数据相比,97%-100%的青藏高原冰川环境微生物为潜在新种;并且大多数微生物主要分布在单一冰川,不同冰川有不同的特有种,这说明了青藏高原冰川微生物空间异质性,全球变暖导致冰川的快速消融将会加速青藏高原冰川微生物多样性的损失。
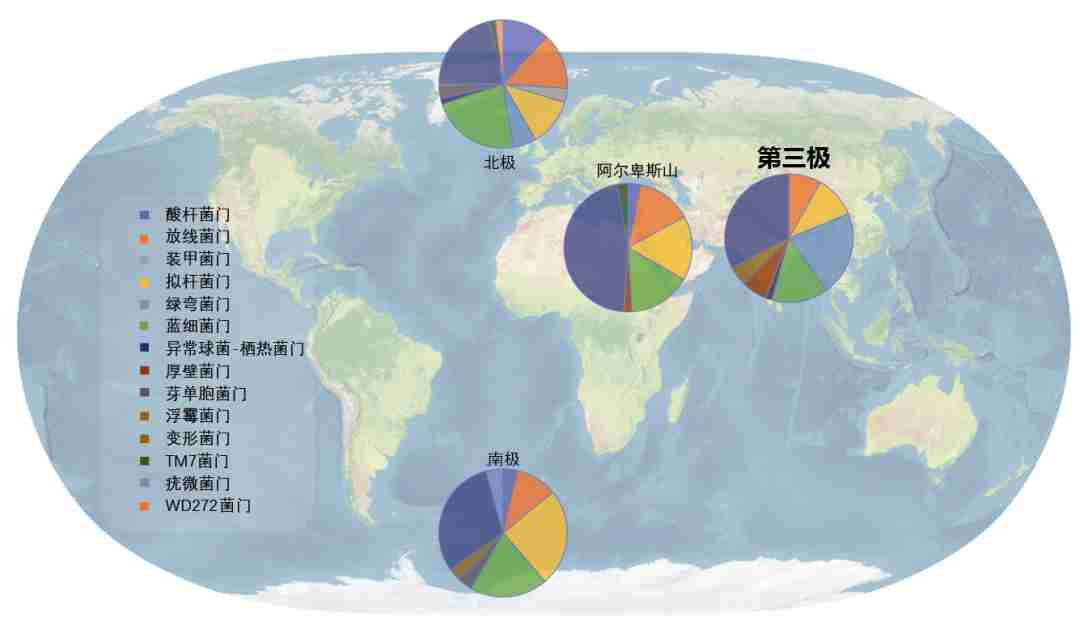
图9.青藏高原冰川微生物群落与其他地区冰川显著不同,图源:第二次科考队。
青藏高原冰川微生物与全球其他冰川微生物显著不同。这种特殊性可能由于青藏高原是同时处于西风和季风作用下,冰川微生物来源与全球其他冰川不同,导致不同类群微生物定殖于青藏高原冰川,进一步说明青藏高原冰川生态系统的生物特殊性。
“冰川来客”有望造福人类健康
青藏高原冰川具有大量待开发可利用的微生物资源,目前已获取青藏高原冰川微生物2532万个基因,其中65%可被注释,剩余35%的功能未知,说明青藏高原冰川微生物基因具有独特性和新颖性,可能编码耐低温和耐紫外辐射功能的新型蛋白。
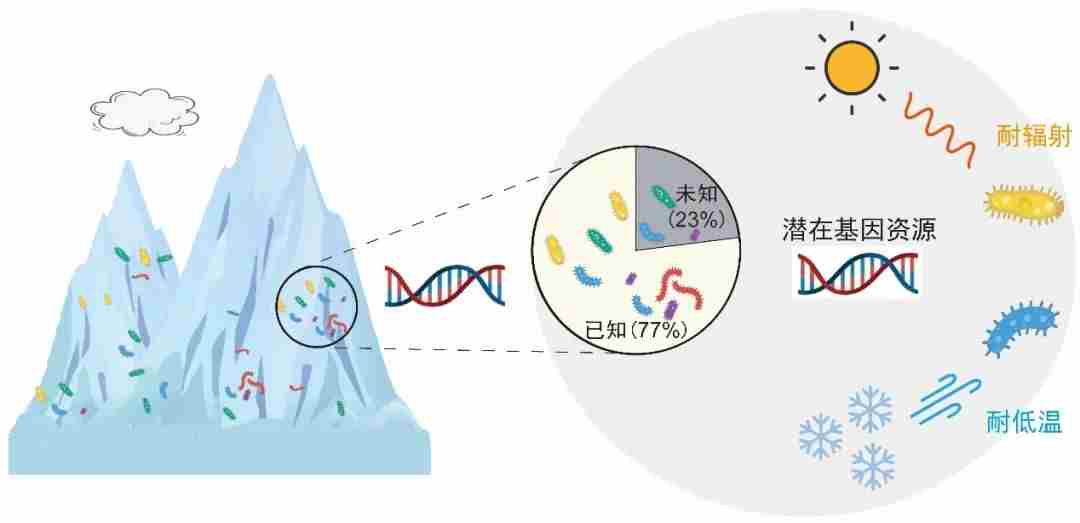
图10.青藏高原冰川蕴藏着独特的微生物,具耐低温和耐紫外辐射功能潜力,图源:第二次科考队。
在所获得的青藏高原微生物基因中,约10%与次级代谢产物合成相关。次级代谢产物不直接涉及到生命正常生长、发育或繁殖,但对生物之间的相互作用和环境适应性具有重要作用。许多次级代谢产物在医药、农业等领域起着重要的作用。冰川微生物的次级代谢产物中,非核糖体多肽和聚酮合成相关的基因占比最高,达31%,其次是与色素相关的萜类合成基因,占26%。非核糖体多肽合成酶及聚酮合成酶可合成多种重要抗生素、抗真菌剂、抗癌和抗肿瘤药物分子,是当前药物领域研究的热点;而萜类合成相关基因可能与冰川微生物抵御紫外辐射、捕获光能等需求有关。
冰川次级代谢产物合成基因展现了强大的利用价值,如一个来自气单胞菌、由近5万个氨基酸组成的非核糖体多肽合成酶,具有合成抗肿瘤能力小分子化合物的潜力;另一个来自黄杆菌、由3.6万个氨基酸组成的非核糖体多肽合成酶,具有合成抗生素的潜力。更多次级代谢产物的功能、生物活性,以及微生物资源在其他领域的应用有待进一步挖掘。
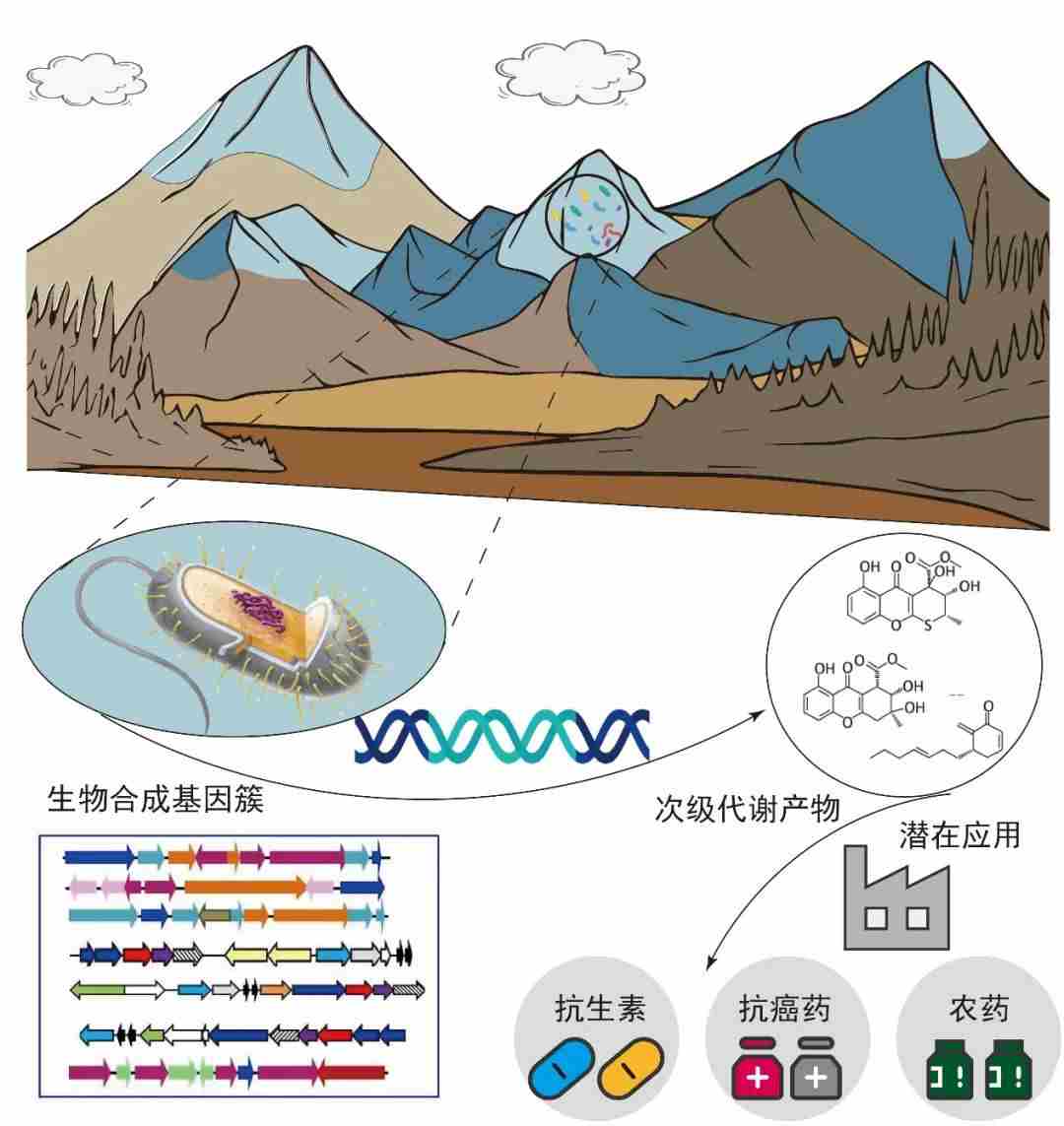
图11.青藏高原冰川微生物次级代谢产物的应用价值,图源:第二次科考队。
获取青藏高原冰川微生物基因组和基因数据是理解冰川生态系统微生物适应机制、碳氮循环过程、生物资源开发和对下游生态影响评估的基础。
冰川消融亦或打开潘多拉魔盒,释放威胁人类健康的有害微生物
气候变暖和人类活动增强导致青藏高原特有微生物损失加速,同时也导致人类相关的有害微生物和相关基因如病毒、致病菌、抗生素抗性基因及毒力因子等向冰川的输送增强,之后伴随圈层之间的相互作用在全球扩散。气候变暖有可能导致“封存”于青藏高原冰川及冻土的远古微生物得以“解封”,对人类生命安全和生态系统健康形成潜在的挑战。如2005年美国国家航空航天局复苏了阿拉斯加冻土中3.2万年前的细菌;2016年西伯利亚一名男孩因感染了75年前死亡的驯鹿尸体中的炭疽而死亡;2022年法国科学家复活了在西伯利亚永久冻土中冷冻了4.85万年的病毒,该病毒复活后仍能感染活细胞,属于潘多拉病毒属(Pandoravirus),是迄今复活的最古老病毒。
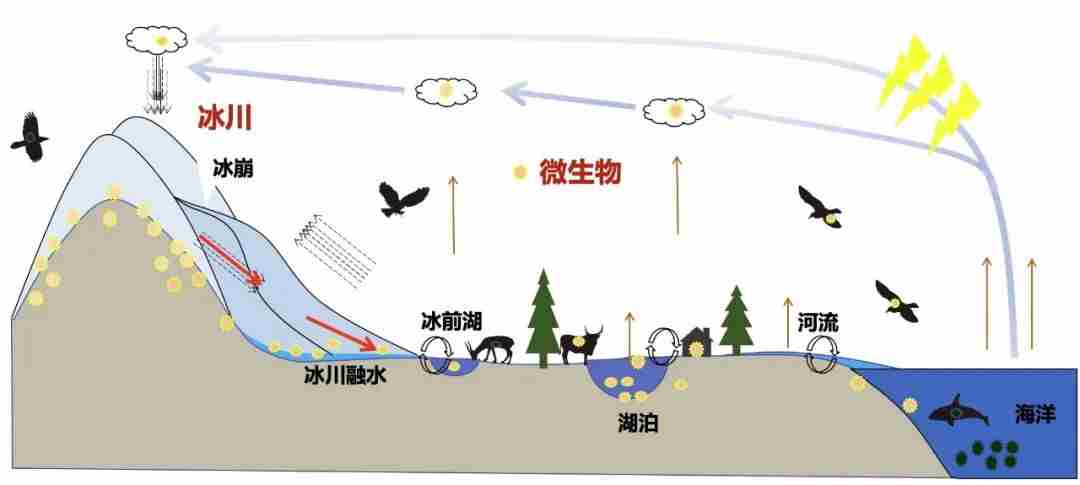
图12.微生物在大气圈-冰冻圈-水圈-生物圈间传播示意图,图源:第二次科考队。
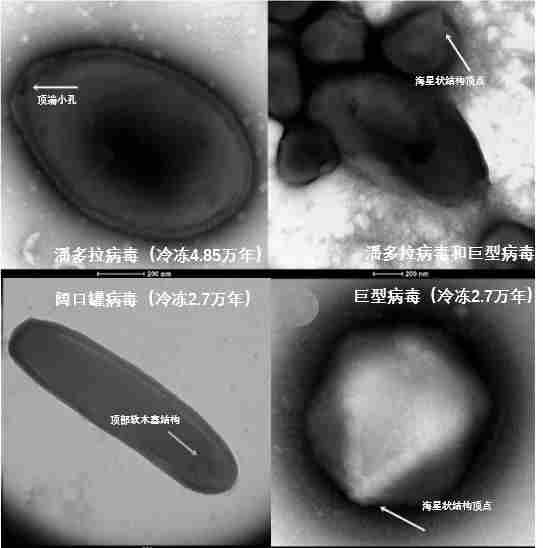
图13.科学家从西伯利亚永久冻土中复活的冷冻了数万年的病毒,图源:引自文献[17]。
2021年,美国俄亥俄州立大学伯德极地和气候研究中心报道,在青藏高原古里雅冰芯中发现了33个病毒物种,其中仅有12%为已知病毒,但它们主要为侵染细菌的噬菌体且数量极低。最近,兰州大学泛第三极环境中心团队研究了青藏高原26条冰川雪、冰和冰尘中的病毒,共发现了8894个病毒物种,种类数远低于土壤、湖泊和海洋等环境,其中西风区冰川中病毒数量大于季风区。尽管38%的病毒为新病毒种类,但以噬菌体为主,对人类无害。与人类相关病毒的相对丰度极低,仅百万分之一,且均为与人类长期共存、被人类熟知的病毒。因此,青藏高原冰川中的病毒对人类社会造成危害的风险极低。
在青藏高原冰川中,虽然与人类相关的病毒风险低,但是科考队员发现了微生物抗生素抗性基因的潜在危害。微生物利用其抗性基因,通过外排、解毒和靶点修饰等机制对抗抗生素,从而有利于微生物自身更好地适应环境,尽管微生物抗性基因的产生是一种自然和古老的过程。但在抗生素大量使用、环境浓度升高的背景下,这些抗性基因可通过水平基因转移在微生物间,及环境与微生物转移和传播,造成了耐药菌的滋生和扩散,甚至出现了能耐受几乎所有抗生素的“超级细菌”,导致重大的公共卫生危机。
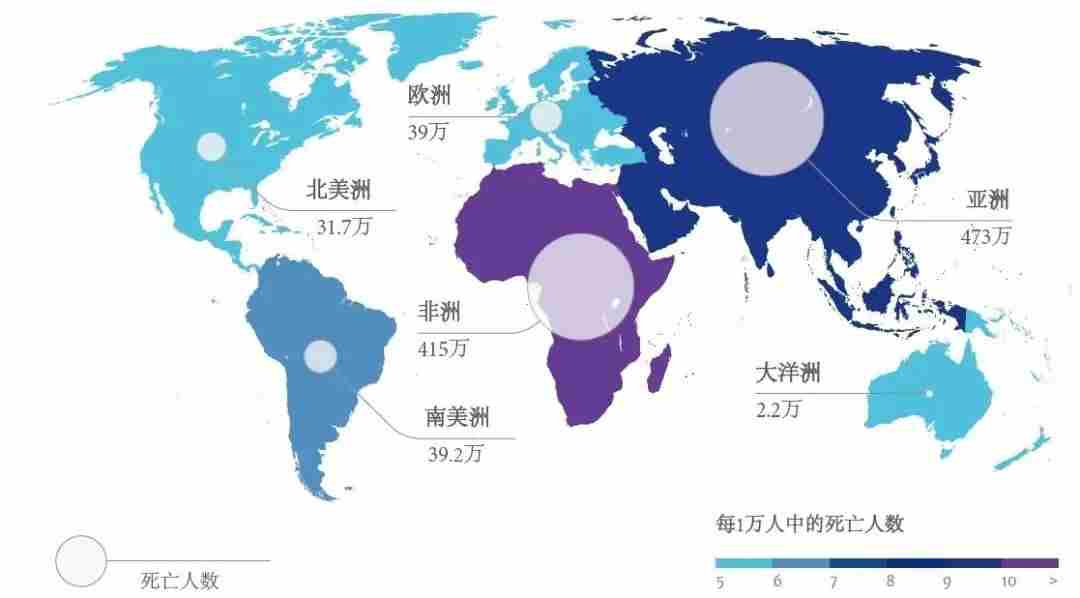
图14.全球预计到2050年每年因耐药性微生物感染致死人数,图源:The Review on Antimicrobial Resistance 2014
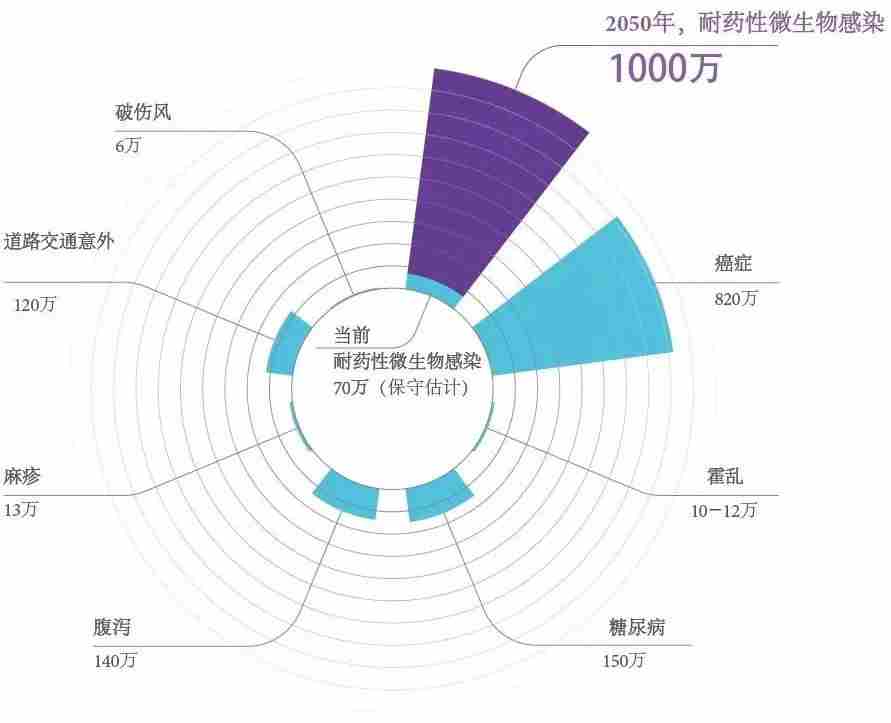
图15.2014年和2050年因耐药性微生物感染致死与其他主要死亡原因死亡人数对比,图源:The Review on Antimicrobial Resistance 2014。
在青藏高原21条冰川的85个样品中,发现了隶属于29个抗性基因大类、145个抗生素抗性基因亚型的抗性基因。抗性基因在季风区冰川雪中的丰度显著高于西风区冰川,季风区冰川降水主要受途经印度及孟加拉等国的印度季风影响,冰川抗性基因丰度与这些国家抗生素的使用量呈显著正相关,表明青藏高原周边国家大量使用抗生素可能导致抗性基因随印度季风传播扩散至青藏高原冰川。此外,从青藏高原冰川样品中分离得到96个携带一种抗性基因的冰川耐药菌,1098个携带两种及以上抗性基因的冰川多重耐药菌,其中有21.04%的冰川耐药菌可划分为致病菌,主要隶属于葡萄球菌属、不动杆菌属和假单胞菌属,被联合国微生物组织列入“极为重要”病原体清单。
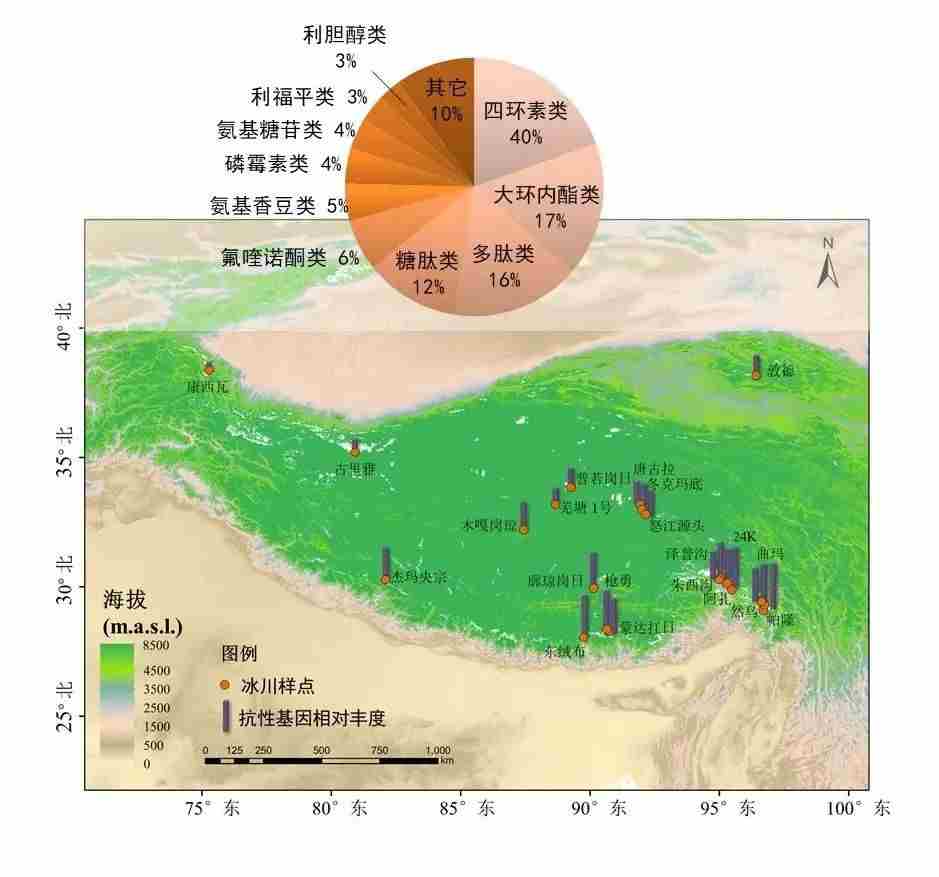
图16.青藏高原21条冰川抗生素抗性基因相对丰度以及主要抗性基因类型,图源:第二次科考队。
气候变暖造成冰川消融,可能会导致耐药菌、耐药致病菌释放到下游生态系统,对下游环境的生态安全和居民健康产生潜在威胁。因此,为发挥青藏高原的生态安全屏障作用,有必要进一步明确青藏高原潜在病原体,特别是人畜共患病的病原体数量和分布格局、致病机制及传播与演化过程。
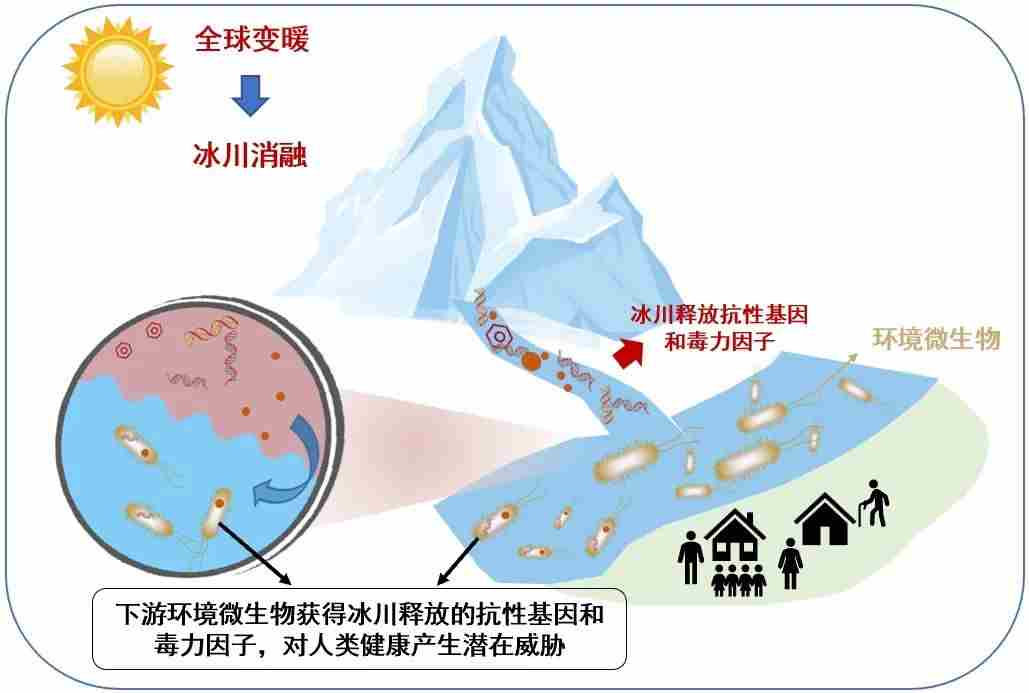
图17.抗性基因和毒力因子向冰川下游传播的示意图,图源:第二次科考队。
微生物在青藏高原大气-冰川-河流-湖泊-土壤间迁移传播,在冰冻圈、水圈、岩石圈多圈层多生境多生态系统中发挥着重要的基石作用,更深入的研究将为评估气候变化对青藏高原温室气体通量的影响提供理论依据、为青藏高原生物资源开发和生态文明建设提供理论依据和数据支撑,进一步增强青藏高原的生态屏障作用,助力青藏高原生态文明建设。
参考文献
[1] Whittaker, R. H. 1969. New concepts of kingdoms or organisms. Evolutionary relations are better represented by new classifications than by the traditional two kingdoms. Science, 163(3863):150-160.
[2] Woese, C. R. and Fox, G. E. 1977. Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proceedings of the National Academy of Sciences of the United States of America, 74(11):5088-5090.
[3] Shu, W.-S. and Huang, L.-N. 2022. Microbial diversity in extreme environments. Nature Reviews Microbiology, 20(4):219-235.
[4] Priscu, J. C., et al. 2007. Biological material in ice cores. Encyclopedia of Quaternary sciences, 2:1156-1166.
[5] Rogers, S. O., et al. 2004. Recycling of pathogenic microbes through survival in ice. Medical Hypotheses, 63(5):773-777.
[6] 刘时银 等. 2015. 基于第二次冰川编目的中国冰川现状. 地理学报, 70(01):3-16.
[7] 姚檀栋 等. 2019. “亚洲水塔”变化与影响. 中国科学院院刊, 34(11):1203-1209.
[8] Xiang, S. R., et al. 2005. Vertical quantitative and dominant population distribution of the bacteria isolated from the Muztagata ice core. Science in China Series D-Earth Sciences, 48(10):1728-1739.
[9] Yao, T., et al. 2012. Different glacier status with atmospheric circulations in Tibetan Plateau and surroundings. Nature Climate Change, 2(9):663-667.
[10] Anesio, A. M., et al. 2009. High microbial activity on glaciers: importance to the global carbon cycle. Global Change Biology, 15(4):955-960.
[11] Anesio, A. M., et al. 2017. The microbiome of glaciers and ice sheets. NPJ Biofilms Microbiomes, 3:10.
[12] Anesio, A. M. and Laybourn-Parry, J. 2012. Glaciers and ice sheets as a biome. Trends in Ecology and Evolution, 27(4):219-225.
[13] 陈德亮 等. 2015. 青藏高原环境变化科学评估: 过去、现在与未来. 科学通报, 60(32):3025-3035.
[14] Liu, Y., et al. 2022. A genome and gene catalog of glacier microbiomes. Nature Biotechnology, 40(9):1330-1331.
[15] Revich, B. A. and Podolnaya, M. A. 2011. Thawing of permafrost may disturb historic cattle burial grounds in East Siberia. Global Health Action, 4.
[16]https://www.natureworldnews.com/articles/45267/20210222/prehistoric-animal-viruses-permafrost-caught-interest-russian- scientists.htm, Nature World News, feb 22, 2021.
[17]Alempic,J.M., et al. 2023. An update on eukaryotic viruses revived from ancient permafrost. bioRxiv, 2022.2011.2010.515937.
[18] Zhong, Z. P., et al. 2021. Glacier ice archives nearly 15,000-year-old microbes and phages. Microbiome, 9(1):160.
[19] 朱永官 等. 2015. 抗生素耐药性的来源与控制对策. 中国科学院院刊, 30(4):509-516.
[20] Wright, G. D. 2007. The antibiotic resistome: the nexus of chemical and genetic diversity. Nature Reviews Microbiology, 5(3):175-186.
[21] Liu, Y. Y., et al. 2016. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infectious Diseases, 16(2):161-168.
[22] Mao, G., et al. 2023. Monsoon affects the distribution of antibiotic resistome in Tibetan glaciers. Environmental Pollution, 317.
第三极大本营
创作团队
策划 |姚檀栋、安宝晟、刘勇勤
姚汝桢、王伟财、戴玉凤
撰文 |刘勇勤、张蔚珍、张志好、计慕侃、刘鹏飞
审核 | 姚檀栋、刘勇勤
编辑 | 戴玉凤、姚汝桢、陈文锋
中国青藏高原研究会
第二次青藏高原综合科学考察研究队
共同主办
旨在连接从智库到公众的知识纽带,
构建原创科普内容的分享平台,
传播内容丰富通俗易懂的科学。
最新科研成果
-
 发布者:
发布者:
世代栖息在“世界屋脊”的裂腹鱼类
-
发布者: 生物多样性保护与可持续利用
青海省野外科考日志3——与鸟类同行
-
 发布者: 生物多样性保护与可持续利用
发布者: 生物多样性保护与可持续利用
青海省野外科考日志2——高寒稀氧全不畏,更展风华向大地